Search
Beranda
Layanan
Dokter
Registrasi
Testimoni
Kontak
Lokasi
Beranda
Layanan
Dokter
Registrasi
Testimoni
Kontak
Lokasi
Berita Kesehatan
Posted by
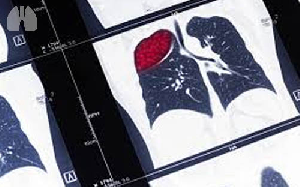
Image :
Kamis, 25 Apr 2024 13:10:59
Sumber :
Berita Kesehatan Lainnya
‹
5 Jenis Operasi yang Paling Sering Direkomendasikan Dokter untuk Kanker Tenggorokan
Penyebab Paru-paru Basah Dipicu oleh Gaya Hidup Tak Sehat
Hipertrofi Konka
Granuloma Paru
Menjaga Kesehatan Gigi Bagi Perokok
Kapas atau Cotton Bud Tersangkut di Telinga? Begini Cara Mengeluarkannya
Tips Memulai Olahraga Supaya Bisa Berhenti Merokok
›
 Copyright ©2017 Klinik Respirasi Malang, All Rights Reserved
Copyright ©2017 Klinik Respirasi Malang, All Rights Reserved